

Sypialnia to nie tylko arena behawioralna — to ekstremalnie dynamiczne pole neurochemiczne, na którym krzyżują się szlaki katecholaminergiczne i serotoninergiczne, determinując przebieg pożądania, ekscytacji, orgazmu oraz fazę refrakcji. Zrozumienie tego układu wymaga porzucenia popularnonaukowych uproszczeń typu „dopamina to przyjemność” i wejścia na poziom molekularny, na którym enzymy limitujące szybkość (rate-limiting enzymes), kofaktory witaminowe oraz antagonizm receptorowy kształtują każdą fazę cyklu seksualnego. Niniejszy tekst rozkłada na czynniki pierwsze układ nagrody oraz chemiczny balans między pożądaniem (dopamina) a ukojeniem (serotonina i oksytocyna), ze szczególnym uwzględnieniem szlaków syntezy, mechanizmów habituacji oraz aminokwasowych prekursorów determinujących dostępność neuroprzekaźników w szczelinie synaptycznej.
Dopamina jako molekuła przewidywania i motywacji
Dopamina nie jest molekułą przyjemności w sensie hedonicznym — jest molekułą przewidywania nagrody (reward prediction error). Model Wolframa Schulza, oparty na rejestracji aktywności neuronów dopaminergicznych w brzusznym polu nakrywki (VTA — ventral tegmental area), wykazał, że te neurony nie odpowiadają na samą nagrodę, lecz na różnicę między oczekiwaną a otrzymaną nagrodą. Gdy nagroda przewyższa oczekiwania — następuje fazowa aktywacja (burst firing) neuronów dopaminergicznych; gdy jest zgodna z oczekiwaniami — brak odpowiedzi; gdy poniżej — następuje tłumienie (pause in firing). To fundamentalne odkrycie przesuwa paradygmat: dopamina koduje nie sam akt seksualny, lecz anticypację, podniecenie wywołane prognozą zbliżającej się nagrody seksualnej.
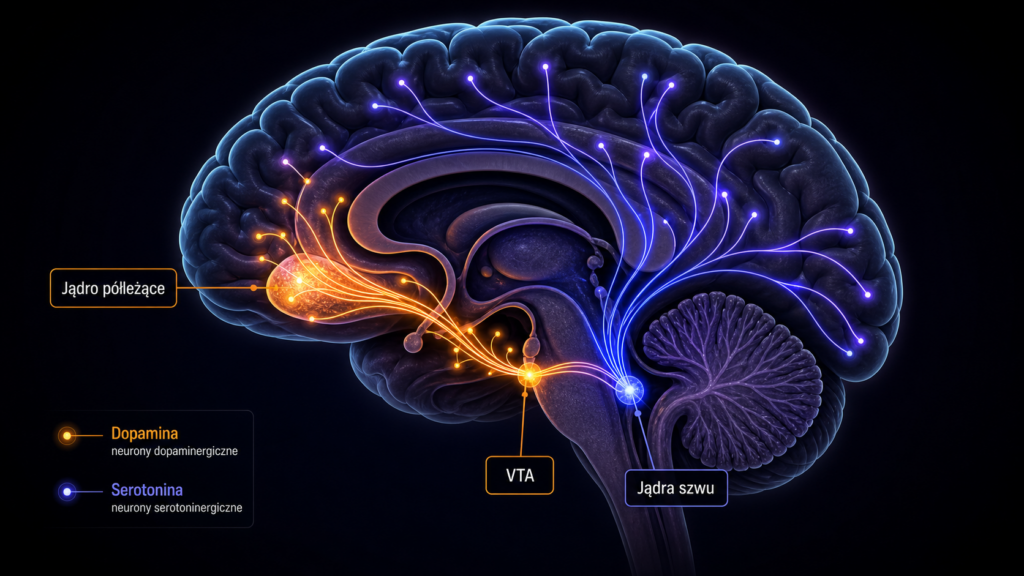
W kontekście behawioru seksualnego dopamina działa poprzez co najmniej trzy integracyjne ośrodki: układ nigrostriatalny (aktywność somatomotoryczna), układ mezolimbiczny (motywacja i salience) oraz przywzrokowe pole środkowe (MPOA — medial preoptic area), który integruje bodźce hormonalne i sensoryczne z pędem kopulacyjnym. To właśnie mezolimbiczny szlak dopaminergiczny — projekcja z VTA do jądra półleżącego (nucleus accumbens, NAc) — stanowi centralny mechanizm motywacyjny nie tylko w kontekście seksualnym, ale też w uzależnieniach od substancji psychoaktywnych.
Szlak mezolimbiczny a fizjologiczny proces budowania napięcia seksualnego
Szlak mezolimbiczny składa się z projekcji dopaminergicznych z brzusznego pola nakrywki (VTA) do jądra półleżącego (NAc). Neurony te znajdują się pod toniczną inhibicją wywieraną przez lokalne interneurony GABA-ergiczne, które z kolei są modulowane przez ligandy receptora opioidowego μ (MOR). Badania immunohistochemiczne z użyciem markera internalizacji MOR wykazały istotny wzrost internalizacji receptorów μ w VTA po kopulacji oraz po samej ekspozycji na bodźce związane z seksem (sex-related environmental cues) — co oznacza, że układ mezolimbiczny aktywuje się endogennie już na etapie antycypacji, przed jakimkolwiek kontaktem fizycznym.
Fizjologiczny proces budowania napięcia seksualnego przebiega następująco na poziomie molekularnym:
- Bodziec seksualny (wzrokowy, olfaktoryczny, dotykowy) aktywuje szlaki sensoryczne docierające do kory przedczołowej i ciała migdałowatego.
- Kora przedczołowa i ciało migdałowate wysyłają projekcje glutaminianergiczne do VTA, znosząc inhibicję GABA-ergiczną.
- Następuje burst firing neuronów dopaminergicznych → uwalnianie dopaminy w NAc.
- Dopamina wiąże się z receptorami D1 (pobudzające,Gs) i D2 (hamujące, Gi) w NAc, modulując wyjście motoryczne i motywacyjne.
- Równolegle MPOA integruje sygnały z estrogenów i androgenów, potęgując pęd kopulacyjny.
Kluczowe jest zjawisko deltaFosB — transkrypcyjnego czynnika akumulującego się w NAc w odpowiedzi na chroniczną stymulację nagrody. Badania na modelach zwierzęcych wykazały, że doświadczenie seksualne powoduje wzrost poziomu deltaFosB w NAc, co prowadzi do strukturalnej neuroplastyczności: zwiększenia liczby kolców dendrytycznych (dendritic spines) oraz sensytyzacji amfetaminowej nagrody. Blokowanie aktywności deltaFosB w NAc za pomocą transferu genów znosi długotrwałe efekty doświadczenia seksualnego — co dowodzi, że deltaFosB jest molekularnym przełącznikiem konsolidacji behawioru nagrody. Co więcej, akumulacja ta oraz sensytyzacja nagrody lekowej zależą od aktywacji receptora D1 (ale nie D2) w trakcie kopulacji.
Zjawisko habituacji — dlaczego nowość podnosi poziom dopaminy i jak zapobiegać rutynie z perspektywy neurobiologicznej
Habituacja to zmniejszenie odpowiedzi neuronalnej na powtarzający się bodziec o stałych parametrach. W kontekście seksualnym manifestuje się to jako stopniowy spadek wyrzutu dopaminy w NAc przy powtarzanej ekspozycji na tego samego partnera lub te same bodźce erotyczne. Klasyczną ilustracją tego mechanizmu jest efekt Coolidge’a — wzrost odpowiedzi seksualnej i skrócenie okresu refrakcji po ekspozycji na nowego partnera.
Badania Fiorino, Coury’ego i Phillipsa (1997) z użyciem mikrodializy in vivo wykazały dynamiczne zmiany wyrzutu dopaminy w jądrze półleżącym podczas efektu Coolidge’a u samców szczurów: po wysyceniu z jedną samicą i spadku dopaminy do poziomu bazalnego, ekspozycja na nową samicę powodowała ponowny, gwałtowny wzrost wyrzutu dopaminy w NAc. Mechanizm ten opiera się na adaptacji receptorowej oraz na kodowaniu predykcyjnym:
- Neurony dopaminergiczne uczą się przewidywać nagrodę na podstawie konkretnych sygnałów kontekstowych (partner, otoczenie, zapach).
- Gdy nagroda staje się w pełni przewidywalna (prediction error = 0), odpowiedź dopaminergiczna maleje.
- Nowość narusza model predykcyjny → ponowny burst firing → nowy wyrzut dopaminy.
Z perspektywy neurobiologicznej zapobieganie rutynie nie polega na eskalacji intensywności bodźców (co prowadzi do tolerancji i potencjalnej patologii), lecz na kontrolowanym wprowadzaniu elementów nowości kontekstowej:
- Zmiana otoczenia i kontekstu sensorycznego (modyfikacja bodźców wzrokowych, olfaktorycznych, dotykowych) resetuje model predykcyjny, generując nowy sygnał błędu predykcyjnego.
- Variability wTiming — zmienność czasowa zbliżeń — zapobiega adaptacji receptorów D1/D2 w NAc.
- Osoby z kompulsywnym zachowaniem seksualnym (CSB) wykazują dysfunkcyjnie nasiloną preferencję nowości seksualnej, pośredniczoną przez większą habituację zakrętu obręczy (cingulate cortex) — co ilustruje, że poszukiwanie nowości ma punkt krytyczny, po którym przeradza się w patologię układu nagrody.
Serotonina i oksytocyna — neurochemia orgazmu i przywiązania
Gdy dopamina buduje napięcie i motywację, serotonina (5-hydroksytryptamina, 5-HT) i oksytocyna wchodzą w fazę rozluźnienia i wiązania. Serotonina jest syntetyzowana w jądrach szwu (raphe nuclei) pnia mózgu i projektuje niemal do całego układu nerwowego, regulując nastrój, sen, apetyt oraz — krytycznie w tym kontekście — hamującą kontrolę nad zachowaniem seksualnym. Oksytocyna, nonapeptyd syntetyzowany w jądrach przykomorowym i nadwzrokowym podwzgórza, uwalniana przez tylny płat przysadki, jest molekułą przywiązania, zaufania i spójności społecznej, ale także kluczowym modulatorem orgazmu i faz poregazmicznych.
Mechanizm spadku dopaminy i nagłego wyrzutu prolaktyny oraz serotoniny w fazie refrakcji
Faza refrakcji — okres po orgazmie charakteryzujący się spadkiem podniecenia, sennością i przejściową niechęcią do dalszej stymulacji — jest neurochemicznym zjawiskiem o wyraźnej dynamice molekularnej:
- Spadek dopaminy: bezpośrednio po orgazmie następuje zahamowanie aktywności neuronów dopaminergicznych w VTA i spadek stężenia dopaminy w NAc. Mechanizm ten obejmuje aktywację autoreceptorów D2 (presynaptycznych, hamujących) oraz aktywację szlaków GABA-ergicznych wygaszających VTA.
- Wyrzut prolaktyny: orgazm wyzwala nagły, pulsacyjny wyrzut prolaktyny z przedniego płata przysadki. Prolaktyna działa jako silny antagonisty zachowania seksualnego — hamuje wydzielanie GnRH (gonadotropin-releasing hormone), obniża stężenie testosteronu i bezpośrednio tłumi responsywność układu dopaminergicznego. Wyższy poziom prolaktyny koreluje z dłuższą fazą refrakcji; u mężczyzn wyższy wyrzut prolaktyny po orgazmie prognozuje dłuższą okres niechęci do dalszej aktywności seksualnej.
- Wyrzut serotoniny: aktywacja układu serotoninergicznego w fazie poregazmicznej nasila uczucie senności, rozluźnienia i satysfakcji. Serotonina moduluje uwalnianie prolaktyny (5-HT stymuluje wydzielanie prolaktyny przez aktywację receptorów 5-HT2A/2C na komórkach laktotropowych przysadki), tworząc sprzężenie zwrotne: serotonina → prolaktyna → hamowanie dopaminy → nasilenie refrakcji.
- Uwalnianie oksytocyny: orgazm wyzwala masywny wyrzut oksytocyny, która promuje przywiązanie do partnera, redukuje reaktywność na stres (hamowanie osi HPA) i moduluje plastyczność synaptyczną w układzie mezolimbicznym, wzmacniając salience nagrody społecznej. Oksytocyna i dopamina działają synergistycznie w NAc — oksytocyna moduluje uwalnianie dopaminy poprzez interakcję z receptorami w obszarze VTA-NAc.
Warto zauważyć, że efekt Coolidge’a — ponowna gotowość do kopulacji w obecności nowego partnera — wiąże się z omijaniem tego mechanizmu prolaktynowo-serotoninowego: nowość reaktywuje szlak dopaminergiczny na tyle silnie, że znosi hamowanie prolaktynowe, skracając fazę refrakcji.
Antagonizm między wysoką serotoniną a pożądaniem — kontekst SSRI
Jednym z najlepiej udokumentowanych klinicznie zjawisk ilustrujących antagonizm serotonina–dopamina w kontekście seksualnym są uboczne skutki leków z grupy SSRI (selective serotonin reuptake inhibitors). Dysfunkcja seksualna jest częstym efektem ubocznym leczenia SSRI, takimi jak paroksetyna, z istotnym wpływem na przestrzeganie zaleceń terapeutycznych i sukces leczenia.
Mechanizm tego antagonizmu na poziomie molekularnym jest wielowarstwowy:
- Inhibycja hydroksylazy tyrozynowej (TH): badania na modelach zwierzęcych wykazały, że paroksetyna (ale nie agomelatonina) powoduje inhibicję ekspresji i aktywności hydroksylazy tyrozynowej w neuronach dopaminergicznych — enzymu limitującego szybkość syntezy dopaminy. Oznacza to, że aktywacja serotoninergiczna bezpośrednio obniża zdolność syntezy dopaminy na poziomie prekursora, nie tylko na poziomie receptorowym.
- Hamowanie zachowań seksualnych przez aktywację receptorów 5-HT2A/2C: serotonina działając przez te receptory w MPOA i NAc hamuje inicjację i utrzymanie zachowania seksualnego. Bupropion, inhibitor wychwytu zwrotnego dopaminy i noradrenaliny (DNRI), przeciwdziała tym efektom i jest stosowany klinicznie w leczeniu dysfunkcji seksualnej indukowanej SSRI.
- Przesunięcie równowagi dopamina/serotonina: wysoka serotoninergia toniczna przesuwa balans w stronę układu hamowania (state satisfaction) kosztem układu dążenia (seeking system), co fenomenologicznie objawia się jako spadek libido, opóźniony orgazm (delayed ejaculation) i anhedonia seksualna.
Z perspektywy ewolucyjnej ten antagonizm ma sens adaptacyjny: serotonina sygnalizuje „status quo zaspokojony” — układ w stanie nasycenia, w którym poszukiwanie nowych zasobów (w tym partnerów) jest nieadaptacyjne. Dopamina sygnalizuje „deficyt / szansa” — układ w stanie poszukiwania, w którym działanie eksploatacyjne jest uzasadnione.
Prekursory aminokwasowe wspierające szlaki neuroprzekaźników
Dostępność neuroprzekaźników w szczelinie synaptycznej jest w pierwszej kolejności limitowana przez dostępność ich prekursorów aminokwasowych oraz aktywność enzymów limitujących szybkość (rate-limiting enzymes). W przypadku dopaminy prekursorem jest L-tyrozyna, a w przypadku serotoniny — L-tryptofan. Zrozumienie kinetyki tych szlaków wymaga analizy nie tylko samej syntezy liniowej, ale też metabolitów krzyżowych i rywalizacji o kofaktory.
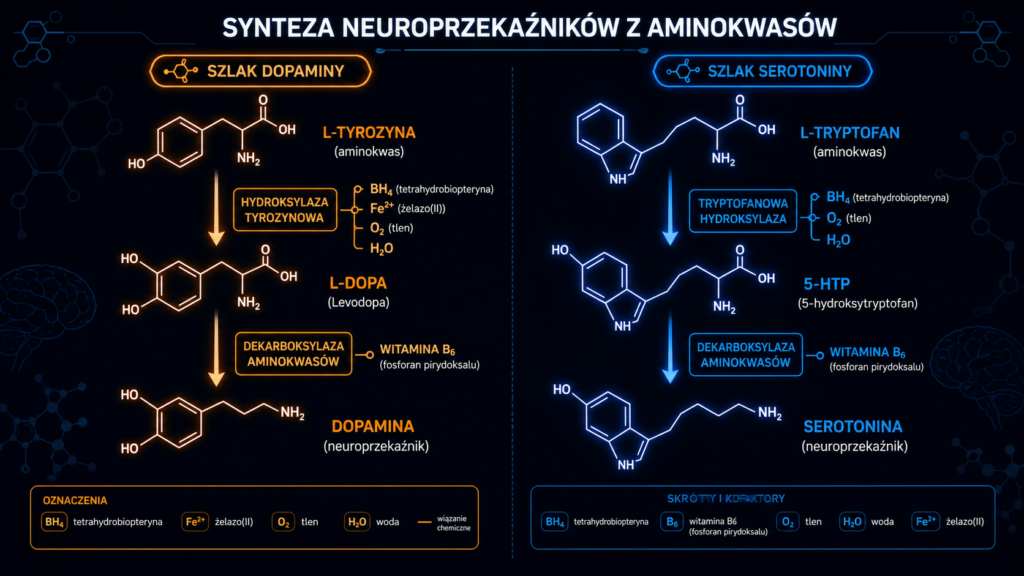
Szlak L-tyrozyny → L-DOPA → dopamina
Synteza dopaminy przebiega dwuetapowo:
- L-Tyrozyna → L-DOPA — katalizowana przez hydroksylazę tyrozynową (TH, EC 1.14.16.2). Jest to enzym limitujący szybkość (rate-limiting enzyme) całego szlaku katecholaminergicznego. TH wymaga tetrahydrobiopteryny (BH4) jako kofaktora oraz jonów Fe²⁺ w centrum aktywnym. BH4 jest syntetyzowany de novo z GTP (szlak de novo) lub regenerowany z dihydrobiopteryny (BH2) przez dehydrogenazę dihydropterydynową (DHPR) z wykorzystaniem NADPH. Niedobór BH4 (np. w niedoborze kwasu foliowego lub mutacjach GCH1) bezpośrednio limituje syntezę dopaminy.
- L-DOPA → Dopamina — katalizowana przez dekarboksylazę aminokwasów aromatycznych (AADC, EC 4.1.1.28), wymagającą fosforanu pirydoksalu (PLP — aktywna forma witaminy B6). Reakcja ta jest szybka i ma wysoką pojemność — dlatego L-DOPA jest stosowana w leczeniu choroby Parkinsona (omija limit TH).
Metabolitami krzyżowymi w szlaku tyrozynowym są:
- Noradrenalina — dopamina jest dalej hydroksylowana przez dopaminową β-hydroksylazę (DBH, wymaga askorbinianu/Cu²⁺) do noradrenaliny, a następnie metylowana do adrenaliny (PNMT, SAM jako donor metylowy).
- Homoproteiny kwasowe — rozpad dopaminy przez MAO (MAO-A i MAO-B) i COMT prowadzi do DOPAC i HVA (kwas homowanilinowy) — markerów metabolizmu dopaminergicznego mierzonych w CSF.
Kluczowy aspekt: L-tyrozyna krzyżuje się ze szlakiem fenyloalaniny — fenyloalanina jest hydroksylowana do tyrozyny przez hydroksylazę fenyloalaninową (również zależną od BH4). Niedobór BH4 blokuje więc obie reakcje, a fenyloketonuria (mutacja PAH) powoduje akumulację fenyloalaniny, która competitywnie hamuje transport innych aminokwasów aromatycznych przez LAT1 (L-type amino acid transporter 1) przez barierę krew-mózg.
Szlak L-tryptofanu → 5-HTP → serotonina
Synteza serotoniny przebiega analogicznie:
- L-Tryptofan → 5-hydroksytryptofan (5-HTP) — katalizowana przez tryptofanową hydroksylazę (TPH, EC 1.14.16.4), również enzym limitujący szybkość, wymagający BH4 i Fe²⁺ jako kofaktorów. Występują dwie izoformy: TPH1 (obwodowo, głównie w komórkach enterochromafinowych jelita) i TPH2 (w ośrodkowym układzie nerwowym, w jądrach szwu).
- 5-HTP → Serotonina (5-HT) — katalizowana przez tę samą dekarboksylazę aminokwasów aromatycznych (AADC, PLP-zależną) co w szlaku dopaminergicznym. To punkt krzyżowy: AADC jest enzymem wspólnym dla obu szlaków, co oznacza, że kompetycja o PLP (witaminę B6) może wpływać na równowagę syntezy dopaminy i serotoniny.
Metabolizm tryptofanu jest jednak znacznie bardziej złożony — tylko około 1% tryptofanu trafia do szlaku serotoninergicznego. Pozostała część ulega metabolizmowi przez trzy rozbieżne drogi:
- Szlak kinureninowy (>95% tryptofanu) — katalizowany przez 2,3-dioksygenazę tryptofanową (TDO) i indoloaminową 2,3-dioksygenazę (IDO). Prowadzi do kinureniny, a następnie do kwasu chinolinowego (agonisty NMDA, neurotoksynnego) lub kwasu kinureninowego (antagonisty NMDA, neuroprotekcyjnego). Aktywacja szlaku kinureninowego (np. w stanie zapalnym, przez interferon-γ i TNF-α) może odciągnąć tryptofan od syntezy serotoniny, prowadząc do hiposerotoninergii — mechanizm ten jest istotny w patofizjologii depresji zapalnej.
- Szlak indolowy mikrobiologiczny — flora jelitowa metabolizuje tryptofan do indolu, który jest dalej siarkowany w wątrobie do siarczanu indoksylu (IS) — czynnika nefrotoksycznego i naczyniowego.
- Szlak melatoninowy — serotonina jest dalej acetylowana (przez AANAT) i metylowana (przez ASMT/HIOMT) do melatoniny w szyszynce. Oznacza to, że szlak serotoninergiczny i szlak melatoninowy współzawodniczą o pulę serotoniny — stąd wysoka aktywność szyszynki w ciemności (aktywacja AANAT przez receptor β-adrenergiczny z nadwzrokowego jądra) może obniżać dostępność serotoniny w innych rejonach mózgu w fazie nocnej.
Kofaktory i rywalizacja o zasoby — perspektywa systemowa
Wspólnymi kofaktorami obu szlaków (dopaminergicznego i serotoninergicznego) są:
- Tetrahydrobiopteryna (BH4) — niezbędna dla TH i TPH; jej synteza i regeneracja determinuje przejściowość syntezy obu neuroprzekaźników.
- Fosforan pirydoksalu (PLP, witamina B6) — kofaktor AADC (wspólny enzym) oraz dekarboksylaz innych aminokwasów; rywalizacja o PLP występuje nie tylko między dopaminą i serotoniną, ale też z syntezą GABA (dekarboksylaza glutaminianowa, GAD).
- Jony Fe²⁺ — kofaktor TH i TPH; niedobór żelaza (szczególnie u kobiet menstruujących) bezpośrednio obniża syntezę obu neuroprzekaźników.
- S-adenozylometionina (SAM) — donor metylowy w metabolizmie dopaminy (COMT) i w syntezie melatoniny z serotoniny; puli SAM determinuje szybkość rozpadu dopaminy oraz konwersji serotoniny do melatoniny.
Transport L-tyrozyny i L-tryptofanu przez barierę krew-mózg zachodzi poprzez ten sam transporter LAT1, co oznacza bezpośrednią kompetycję między tymi aminokwasami o transport do mózgu. Podwyższenie poziomu jednego aminokwasu może obniżyć dostępność drugiego w OUN. Co więcej, inne aminokwasy neutralne o dużych łańcuchach (leucyna, izoleucyna, walina, fenyloalanina) również korzystają z LAT1 — dlatego spożycie białka bogatego w BCAA może rywalizować o transport tryptofanu do mózgu, zmniejszając syntezę serotoniny. Zjawisko to jest podstawą tzw. „paradoksu tryptofanowego” — suplementacja samym tryptofanem może być mniej efektywna w podnoszeniu poziomu serotoniny mózgowej niż sugerowałaby jej biodostępność, z powodu kompetycji transportowej.
Z kolei insulinowy mechanizm tryptofanowy polega na tym, że wyrzut insuliny po spożyciu węglowodanów obniża stężenie aminokwasów neutralnych (ale nie tryptofanu, który jest w dużej mierze związany z albuminą) w osoczu, co zwiększa stosunek tryptofanu do aminokwasów neutralnych (Trp:LNAA) i promuje transport tryptofanu do mózgu. To neurochemiczne uzasadnienie zjawiska „senności po węglowodanach” — węglowodany, poprzez insulinę, zwiększają dostępność tryptofanu dla syntezy serotoniny (a następnie melatoniny) w mózgu.
Implikacje dla balansu neuroprzekaźników w kontekście seksualnym
Z perspektywy syntezy neuroprzekaźników balans dopamina–serotonina w sypialni jest determinowany przez:
- Dostępność i proporcję prekursorów (tyrozyna vs tryptofan) w diecie oraz ich transport przez BBB.
- Status kofaktorów: BH4, PLP, Fe²⁺, kwas foliowy — ich niedobór obniża syntezę obu neuroprzekaźników.
- Stan zapalny (aktywacja szlaku kinureninowego odciągającego tryptofan od syntezy serotoniny).
- Rytm dobowy (konkurencja o serotoninę między układem serotoninergicznym i melatoninowym w różnych fazach cyklu dobowego).
Z perspektywy motywacji i neurobiologii przyjemności zrozumienie tych szlaków pozwala na świadome modelowanie równowagi między dążeniem (seeking — dopamina) a satysfakcją (liking/safety — serotonina/oksytocyna) — co w kontekście sypialni przekłada się na zdolność do budowania napięcia, doświadczania orgazmu oraz faz poregazmicznego przywiązania.